作物の糖・デンプン代謝に関する生産生理学的研究
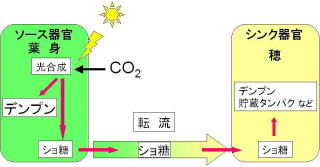 作物において、光合成を行う葉身(ソース)において固定された炭素は糖(イネやダイズではショ糖)に変換された後、収穫部位(シンク)に転流し、デンプンや糖として蓄積します。当研究室ではこのような植物体内での糖・デンプン代謝に着目し、主にイネ、ダイズ、ソルガムを材料とした研究を行っています。
作物において、光合成を行う葉身(ソース)において固定された炭素は糖(イネやダイズではショ糖)に変換された後、収穫部位(シンク)に転流し、デンプンや糖として蓄積します。当研究室ではこのような植物体内での糖・デンプン代謝に着目し、主にイネ、ダイズ、ソルガムを材料とした研究を行っています。
植物の糖・デンプン代謝は糖・デンプンを合成・分解する酵素や糖を輸送する輸送体の働きによって制御されています。当研究室では、各酵素や輸送体およびそれらをコードする遺伝子に着目し、突然変異系統や染色体断片置換系統、準同質遺伝子系統、遺伝子組換え系統といった様々な材料を用いて作物の糖・デンプン代謝の制御機構を明らかにするとともに、遺伝資源として有用な系統の選抜・解析を行っています。
また、飼料用イネやスイートソルガムといった作物は茎部に糖を多く蓄積するため、バイオエタノールの原料や家畜の飼料用として有用ですが、その糖蓄積のメカニズムは明らかとなっておりません。そこで当研究室では食用イネと飼料用イネ、食用であるグレインソルガムと飼料・バイオエタノール用であるスイートソルガムを糖・デンプン代謝の観点から比較することで、糖蓄積機構を明らかにしようとしています。
以上のような糖・デンプン代謝に着目した解析を通じて、作物の収量向上および、茎部高糖性品種の糖蓄積機構の解明を目指しています。
主な研究成果
・イネのショ糖リン酸合成酵素(SPS)に関する機能解析
イネにおいて、光合成によって固定された炭素はショ糖(スクロース)としてシンク器官へと転流し、植物体の成長や子実の生産に利用されます。植物におけるショ糖合成系の鍵酵素の1つとしてショ糖リン酸合成酵素(Sucrose
Phosphate Synthase, SPS)が知られており、様々な作物でSPSが転流効率の制御を通じて生育、収量形成に重要な役割を果たすことが明らかとなっています。
イネのSPSは5つの遺伝子(SPS1, 2, 6, 8, 11)によってコードされていますが、それらの遺伝子の発現特性や生理的な分業は明らかとなっていませんでした。当研究室の解析から、5つのSPS遺伝子はすべて複数の器官で発現している一方で、遺伝子によって発現の高い器官が異なっており、生理的な分業を行っていることが明らかとなりました。また、遺伝子によって発現制御機構が異なる可能性が示唆されました(Okamura
et al., 2011)。そこで、次にSPS1とSPS11に着目し、生物発光リアルタイム測定法を用いてプロモーター活性を調べたところ、SPS1とSPS11は光と概日時計によって制御されている一方、ショ糖によっては制御されていないことが明らかとなりました(Yonekura
et al., 2013)。
次に、葉身で主に発現しているSPS1に着目しました。コシヒカリを遺伝的背景としSPS1遺伝子近傍がインディカ品種カサラスに置き換わった準同質遺伝子系統NIL-SPS1の解析から、SPS1が一穂あたりの籾数の制御に関与している可能性が示唆されました(Hashida
et al., 2013)。また、SPS1遺伝子破壊系統の解析から、SPS1が花粉の発芽に必須であることが示されました(Hirose et al.,
2014)。
以上の研究からSPS遺伝子の発現特性、発現制御機構の一端が明らかになりました。また、生育や収量形成における多面的な役割も明らかになりつつあります。今後はSPSがイネの生育や収量にどのような役割を果たしているか、各遺伝子の発現特性の違いにも着目しながら明らかにしていきたいと考えています。
・イネの茎部におけるデンプン蓄積の意義
イネは光合成で固定された炭素のうち、呼吸や成長に使われなかった余剰分を主にデンプンの形で茎に蓄積します。このデンプンは出穂後に穂に転流し、種子登熟の貴重な炭素源となります。特にその貢献度は寡照や低窒素などの不良条件下で高まるとされていることから、イネの茎部におけるデンプンの出穂前蓄積は、収量の増加や安定化に大きく関わっていると考えられています。
そこで当研究室では、デンプン合成の鍵酵素として知られているADP グルコースピロホスホリラーゼ(ADP-glucose pyrophosphorylase,
AGPase)をコードする遺伝子のうち、茎部特異的に発現する遺伝子である、AGPL1遺伝子破壊系統を用いて、イネの茎部におけるデンプン蓄積の生理・生態学的意義を解析しました。
その結果、AGPL1遺伝子破壊系統では出穂期の茎のデンプン濃度が大きく低下し、子実収量は野生型系統よりも4%ほど低下しました(Okamura
et al., 2013)。この収量の低下程度は、登熟中に遮光幕により日射量を減らすことでさらに大きくなり、23%にも昇りました。このことから、茎部におけるデンプンの蓄積が寡照条件下における子実の十分な登熟に必要不可欠であることが直接的に示されました。
さらに当研究室は、AGPL1遺伝子破壊系統は分げつの角度が拡がった形質を示しており、茎の重力に対する応答性も低くなっていることを発見しました(Okamura
et al., 2013; 2015)。植物においてデンプンを蓄積したアミロプラストは重力の方向を感知する平衡石のような役割を担っていることが知られているため、AGPL1遺伝子破壊系統では茎のデンプン蓄積量の低下により重力への応答性が悪くなり、その結果分げつ茎が垂直方向に立ちにくくなったため、分げつ角度が拡がった草型を示したと考えられました。このように本研究により、イネの茎部におけるデンプン蓄積は、不良条件下における登熟に利用される炭水化物の予備的な蓄積としての機能に加えて、分げつの角度などの草型を制御する上でも重要な役割を担っていることが明らかになりました。

東京大学農学部 作物学研究室
〒113-8657
東京都文京区弥生1-1-1
Tel:03-5841-5193
Fax:03-5841-8048
Last Update : 2016.4.27